An increase in monocytes in the analysis causes concern for patients. Experienced doctors know that the content of only one type of blood cell cannot make any conclusion about the state of health. There are no clear answers to the question of why some cells are increased and others are decreased.
Any changes in blood tests are used as an addition to the symptoms of the disease and are taken into account in differential diagnosis and treatment.
To understand when and in what ways elevated levels of monocytes cause pathology in the body, we need to remember the role of these cells in supporting health.
Basic functions of monocytes
Increased level of monocytes in the blood
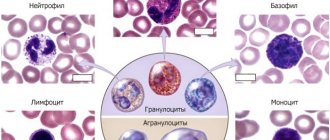
Monocytes are single-celled mature leukocytes, formed in the bone marrow, their life expectancy does not exceed three days. In the bloodstream they reach their most active state. Here they grow, function, and after about 70 hours they degenerate into macrophages, after which they penetrate into nearby tissues.
Protective and cleansing – this is their main function. Moving along the blood stream, they find foreign bodies such as cancer cells, infections, viruses, parasites and try to destroy them. To do this, the cell gets close to the detected problem, envelops it with its body, neutralizes it and removes it from the body. In the same way, monocytes destroy bacteria, dead cells and other substances that pollute the human body.
Macrophages act on the same principle, but they take longer to destroy harmful cells.
Monocytes also take part in the synthesis of interferon, which makes cells immune to the virus, thereby enhancing the body’s protective functions.
Thus, an increase in the content of monocytes means that there is a disease that the body is fighting hard against by increasing protective cells.
Detailed description of the study
In case of damage to the vascular wall, the blood clotting process is activated. This complex process involves numerous blood clotting factors, which are produced mainly by the liver and cells of the vascular wall (endothelium).
An important point in stopping bleeding is the formation of a “plug”, consisting of platelets and special blood proteins, at the site of injury. The platelets are directed to the damaged area, stick together and cover it. The blood coagulation cascade is activated.
As the cascade nears completion, the soluble fibrinogen protein is transformed into insoluble fibrin strands. These threads join together to form a fibrin mesh, which is attached to the damaged area of the vessel.
To form a stable fibrin clot, there must be a sufficient number of platelets and clotting factors in the body. But at a certain point this process must stop to avoid unnecessary blood clots.
To control the blood clotting process and prevent excessive thrombus formation, the body has the so-called. fibrinolytic system. One of its components is the protein plasminogen.
Plasminogen is a precursor of the protein plasmin. It is synthesized predominantly by hepatocytes. Plasminogen is activated by a number of enzymes and is transformed into plasmin, the main function of which is to dissolve fibrin strands in a fibrin clot. Plasmin also ensures the healing of damaged areas of the mucous membranes and skin, cleaning the surface of the wound from excess fibrin.
Plasminogen is a single chain glycoprotein. Its receptors are located on the surface of various types of cells, such as monocytes, macrophages, platelets, fibroblasts, endothelial, etc.
Violations of fibrinolysis (dissolution of fibrin clots) can provoke thrombosis or bleeding.
Determination of plasminogen activity in blood serum is used to assess the functional activity of the fibrinolytic system of the blood.
Plasminogen activity increases with:
- Acute inflammatory reactions (infections, viruses, injuries);
- Surgical interventions;
- Pregnancy.
Plasminogen activity decreases in liver diseases, disseminated intravascular coagulation (a pathological process characterized by a malfunction of the blood coagulation and anticoagulation systems), as well as with long-term thrombolytic therapy.
Also, in some cases, a person may have congenital (hereditary) plasminogen deficiency.
Normally, plasminogen in pregnant women physiologically increases in each trimester. This is due to changes in the hemostatic system under the influence of hormonal factors and is necessary for adequate blood supply to the fetus.
Studying the level of plasminogen is necessary to assess disorders of the hemostatic system and determine the tactics of further therapy to prevent possible complications.
Assessment/monitoring of the state of the fibrinolytic system is carried out during the planning process and during pregnancy.
How is the level of monocytes in the blood determined?
In order to determine the number of monocytes contained in the blood, it is enough to take a general blood test. Monocytes are a type of leukocyte, calculated by a laboratory assistant using a special leukocyte formula. There are two indicators of monocytes:
- Absolute quantity
– designated as “monocytes abs” or “monocytes mono”, the average number of cells per 1 μl (microliter) is taken into account.
- Relative quantity
– designated as “value” ppm/l, calculated as a percentage
When is it necessary to see a doctor and take an OAC test?
There are no specific signs indicating that monocytes are elevated. But at the same time, monocytosis is accompanied by other diseases, the symptoms of which can indirectly suggest changes in the number of leukocytes.
You should consult a doctor and get tested if:
- Decreased or complete loss of appetite.
- Increased fatigue and causeless weakness.
- Aversion to meat dishes.
- Sudden weight loss.
- Insomnia and drowsiness.
- Irritation, apathy, nervous breakdowns.
- Increased psycho-emotional excitability.
- Loud motility of the gastrointestinal tract.
- Constant feeling of anxiety.
- Unexplained pain in the abdomen.
- Panic attacks.
- The appearance of foamy stools.
- Stool disorders.
- The appearance of pain in joints and muscles.
- Bloody impurities in the stool.
- The appearance of acne on the mucous membranes.
- Skin rashes.
- The presence of a long dry cough with bloody sputum.
- Rashes and redness on the genitals, as well as in case of heavy discharge from the genital tract.
- Discomfort and pain during sexual intercourse.
Deviations and normal values
In the period from birth to the age of 16 years, the norms of monocytes in the blood change. Absolute values generally decrease, in contrast to relative values, which up to 16 years of age can either decrease or increase.
| Person's age | Norm of MON parameters |
| In a newborn | 3-12% |
| First two weeks of life | The indicator increases to 5-12% |
| By the year | Decreases 4-10% |
| By 2 years | Is 3-10% |
| From 3 to 16 years old | The lower limit remains 3%, and the upper limit is reduced to 1% |
| From 16 years to old age | from 3-11% |
Absolute indicators from birth to 16 years should decrease, initially ranging from 1.9-2.4 million/l, ultimately decreasing to 0.004-0.08 million/l
A reduced level of monocytes in the blood is called monocytopenia, an increased level is called monocytosis.
The focus of inflammation and an increased level of monocytes in the transcript will be designated as mono. Absolute monocytosis is an increase in the absolute number of monocytes.
The percentage change in monocytes relative to the total number of leukocytes is called relative monocytosis. From time to time, this indicator may increase as a percentage for various reasons, for example, a very hearty breakfast. That is why minor deviations from the norm have no diagnostic value.
But an increase in white cells in the range of 13-17% may indicate a focus of inflammation, and an indicator of 18-24% indicates a serious infectious-inflammatory process.
Standards
The norms for women and men are practically no different. Determination of the absolute (abs.) value per 1 liter of blood is carried out by general analysis and examination of a stained smear. The content of monocytes relative to the total amount of leukocytes is calculated as a percentage and is called the level.
Both indicators are important to evaluate the result. With a sharp fluctuation in the number of other cells included in the leukocyte formula, the level of monocytes may change (above normal or decrease). Although their absolute value will remain unchanged.
Analysis of the relationship with the age category showed an increased level in children under 6 years of age compared to the level in an adult. You can read more about the norms for the number of monocytes in a child’s blood here.
For adults, the normal absolute value is considered to be from zero to 0.08 x 109/l; for a child, from 0.05 to 1.1 x 109/l is acceptable.
In the leukocyte formula, the percentage of monocytes in children is considered normal - 2-12% after birth, in the first 2 weeks - 5-15%, in adults - 3-11%. A similar indicator during pregnancy does not exceed normal limits:
- first trimester on average 3.9%;
- second - 4.0;
- third - 4.5.
Any indicator exceeding the upper limit is called monocytosis and has its own physiological and pathological causes
Causes of increased monocytes
Absolute monocytosis can develop in an adult in the following cases:
- For chronic intestinal diseases such as Crohn's disease, ulcerative colic, inflammation of the small intestine.
- Severe chronic diseases: rheumatoid or psoriatic arthritis, lupus erythematosus.
- Poisoning with substances containing: phosphorus, chlorine or their compounds.
- Infections with infectious diseases: brucellosis, syphilis, salmonellosis, tuberculosis, chickenpox, dysentery, rubella, influenza, whooping cough.
- Rheumatological diseases.
- Rheumatism, endocarditis.
- Blood diseases: chronic myeloid leukemia, osteomyelofibrosis, polycythemia, thrombocytopenic purpura, acute leukemia.
- The development of sepsis or the presence of purulent foci.
- Penetration of parasites and viruses into the body.
- Injury
- Malignant neoplasms of the lymphatic system: lymphoma, lymphogranulomatosis.
- Myocardial infarction.
- Development of fungal diseases.
Also, an excess of the norm of monocytes is observed in patients during the postoperative period and during the recovery period after infectious diseases. Lack of sleep, stress, a rich meal before taking the test, and excessive physical activity can cause an increase in monocytes. Therefore, it is necessary to take this data into account when taking the OAC.
In men
There are no specific reasons for increasing the level of monocytes in men. Men who often experience stress, work a lot, and work is closely related to mental stress and excessive nerves can suffer from monocytosis.
Among women
Elevated monocytes in a woman’s body indicate the presence of infectious diseases, as well as various inflammatory processes. The norms of leukocyte deviation do not depend on gender, however, in women, unlike men, monocytes can increase due to the characteristics of the reproductive system: during menstruation or ovulation, this is associated with a hormonal surge.
In pregnant women
During pregnancy and lactation, the level of leukocytes is especially important for a woman’s body. This is due to the fact that now the protective function must be performed not only for the matter, but also for the fetus inside it, and after the baby is born, they help the woman regain strength faster. During pregnancy, a restructuring of the immune and endocrine systems occurs in a woman’s body, and hormonal levels often change, so the ratio of all types of leukocytes changes.
The level of lower monocytes in the first trimester of pregnancy drops to 1%. But after just a few weeks it increases again to 3%, this figure remains until the last trimester, in which, due to changes in hormonal levels in connection with the body’s preparation for childbirth, the level of monocytes may increase. Immediately after childbirth, the level drops, but later rises again, which means that the body has begun to recover after childbirth.
In children
The normal levels of monocytes in the blood of children are higher than those of adults; the highest levels are observed in newborns. However, there is nothing to worry about if the age norms are not exceeded or the deviation level is not higher than 10%. An increase in this indicator may indicate the presence of infectious or viral diseases or infection with helminths. Often the reason for an increase in leukocyte levels can be the replacement of baby teeth with molars, this is associated with the formation of new tissues.
Also, an increase in monocytes may be associated with hereditary factors, the postoperative period and the presence of more severe diseases, for example, leukemia. It is worth noting that children often experience a low level of leukocytes (monocytopenia) than monocytosis; this is more dangerous and may indicate exhaustion of the body and weak resistance to harmful factors.
According to the “M1/M2” paradigm, there are two subtypes of activated macrophages—classically activated (M1) and alternatively activated (M2), which express various receptors, cytokines, chemokines, growth factors, and effector molecules. However, recent data indicate that, in response to changes in microenvironmental signals, macrophages may exhibit unique properties that do not allow them to be classified into any of these subtypes.
Macrophages play a major role in the body's response to implanted material - catheters, stents, endoprostheses, dental implants. Macrophages phagocytize wear particles from the surface of joint prostheses, initiate inflammation in the prosthetic area and osteolysis, and control the formation of a fibrous capsule around foreign bodies. A brief overview of the factors causing migration, adhesion and activation of macrophages, analysis of their functional characteristics on various surfaces, including biodegradable and non-degradable materials in vivo and in vitro, is presented.
Introduction
Modern medicine is currently impossible to imagine without the use of implantable products installed in the body for various periods of time in order to restore the anatomy and function of organs and tissues lost or affected by a pathological process. The biocompatibility of synthetic materials or tissue-engineered constructs is a major issue affecting the results of such implantations. The reaction to prosthetic material develops in the following sequence: tissue alteration, infiltration by cells of acute, then chronic inflammation with the formation of granulation tissue and fibrous capsule. The severity of these reactions determines the biocompatibility of the implanted device. Macrophages play a major role in the body’s reaction to the installed material - catheters, stents, endoprostheses, dental implants, etc.
Morphology of macrophages
Macrophages are a heterogeneous cell population. The macrophage has an irregular, stellate, multi-processed shape, folds and microvilli on the cell surface, an abundance of endocytic microvesicles, primary and secondary lysosomes. The round or ellipsoid nucleus is located centrally, heterochromatin is localized under the nuclear envelope. The structural features of a cell largely depend on its organ and tissue affiliation, as well as on its functional status. Thus, Kupffer cells are characterized by a glycocalyx, alveolar macrophages contain lamellar (surfactant) bodies, a well-developed Golgi complex, a rough endoplasmic reticulum and many mitochondria, while in microglial cells there are few mitochondria. In the cytoplasm of peritoneal and alveolar macrophages there is a large number of lipid bodies containing substrates and enzymes for the generation of prostaglandins [1]. Adhering and moving macrophages form short-lived, actin-containing structures - podosomes - in the form of a dense central part with microfilaments radiating from them. Podosomes can fuse to form higher-order structures—rosettes—that effectively destroy proteins in the underlying extracellular matrix[2].
Functions of macrophages
Macrophages phagocytose foreign material and cellular tissue detritus, stimulate and regulate the immune response, induce an inflammatory response, and participate in reparative processes and the exchange of extracellular matrix components. The variety of functions performed explains the expression by these cells of a large number of receptors associated with the plasma membrane, intracellular and secreted. Innate immune receptors PRR (pattern-recognition receptors) are activated by a wide range of ligands (with the exception of CD163), providing recognition of highly conserved structures of most microorganisms, the so-called PAMPs (pathogen-associated molecular patterns, pathogen-associated patterns) and similar with them endogenous molecular structures DAMP (damage-associated molecular patterns), formed as a result of damage and cell death, modification and denaturation of protein structures of the extracellular matrix. Most of them mediate endocytosis and elimination of potentially dangerous endogenous and exogenous agents, but at the same time, many of them perform signaling functions, regulating the synthesis of proinflammatory mediators, promoting adhesion and migration of macrophages (Table) [3–7].
The plasma membrane of monocytes/macrophages also expresses specialized receptors that bind one or more structurally similar ligands: the Fc fragment of immunoglobulin G, growth factors, corticosteroids, chemokines and cytokines, anaphylotoxins and costimulatory molecules. The functions of many of these receptors are mediated not only by the binding of ligands, but also by interaction with other receptors (C5aR-TLR, MARCO-TLR, FcγR-TLR), which ensures fine regulation of the synthesis of pro- and anti-inflammatory mediators [2, 6, 8, 9]. A feature of the macrophage receptor system is the presence of trap receptors for proinflammatory cytokines and chemokines (Il-1R2 on M2a macrophages; CCR2 and CCR5 on M2c macrophages), the activation of which blocks the intracellular transmission of the corresponding proinflammatory signal. The expression of cellular receptors is species-, organ- and tissue-specific and depends on the functional status of macrophages. Macrophage cellular receptors studied in detail are shown in the table.
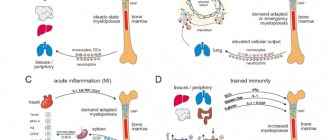
Migration of monocytes/macrophages
Tissue macrophages are derived primarily from blood monocytes, which migrate into tissues and differentiate into different populations. Macrophage migration is directed by chemokines: CCL2 CCL3, CCL4, CCL5, CCL7, CCL8, CCL13, CCL15, CCL19, CXCL10, CXCL12; growth factors VEGF, PDGF, TGF-b; fragments of the complement system; histamine; granule proteins of polymorphonuclear leukocytes (PMNL); phospholipids and their derivatives.
At the initial stages of the inflammatory response, PMNs organize and modify a network of chemokines by secreting CCL3, CCL4 and CCL19 and releasing azurosidine, LL37 protein, cathepsin G, defensins (HNP 1-3) and proteinase 3 preformed into granules, which ensure the adhesion of monocytes to the endothelium, thereby thereby exhibiting the properties of chemoattractants. In addition, PMN granule proteins induce the secretion of chemokines by other cells: azurosidine stimulates the production of CCL3 by macrophages, and proteinase-3 and HNP-1 induce the synthesis of CCL2 by the endothelium. PMN proteinases are capable of activating many protein chemokines and their receptors. Thus, proteolysis of CCL15 by cathepsin G greatly enhances its attractive properties. Apoptotic neutrophils attract monocytes through signals presumably mediated by lysophosphatidylcholine[10].
Any tissue damage leads to the accumulation of macrophages. In the area of vascular injury, the blood clot and platelets release TGF-β, PDGF, CXCL4, leukotriene B4 and IL-1, which have pronounced chemoattractive properties against monocytes/macrophages [11–13]. Damaged tissues are a source of so-called alarmins, which include components of the destroyed extracellular matrix, heat shock proteins, amphoterin, ATP, uric acid, IL-1a, IL-33, mitochondrial DNA of cellular debris, etc. They stimulate the remaining viable cells of damaged tissues and the endothelium of blood vessels to the synthesis of chemokines, some of them are direct factors of chemotaxis [13, 14]. Infection of tissues leads to the appearance of so-called pathogen-associated molecules: lipopolysaccharides, cell wall carbohydrates and bacterial nucleic acids. Their binding by membrane and intracellular receptors of macrophages triggers the process of expression of chemokine genes, which provide additional recruitment of phagocytes [11, 15].
Macrophage activation
Macrophages are activated by a variety of signaling molecules, causing their differentiation into various functional types (Fig. 1). Classically activated macrophages (M1 phenotype) are stimulated by IFNg, as well as IFNg together with LPS and TNF. Their main functions are the destruction of pathogenic microorganisms and the induction of an inflammatory response. Polarization in the M1 direction is accompanied by the secretion of proinflammatory mediators. They express receptors for IL-1 – IL-1R1, TLRs and co-stimulatory molecules, the activation of which ensures amplification of the inflammatory response. Along with pro-inflammatory cytokines, macrophages also secrete the anti-inflammatory cytokine IL-10, with a characteristically high IL-12/IL-10 ratio [11, 13, 16–18]. The bactericidal properties of M1 macrophages are determined by the production of free radicals of nitrogen and oxygen generated by iNOS and the NADPH oxidase complex [19, 20]. Being effector cells in the body's response to bacterial infection, they, at the same time, suppress the adaptive immune response by inhibiting the proliferation of stimulated T cells. IL-12 secreted by M1 macrophages plays a key role in Th1 polarization, and IL-1b and IL-23 direct the immune response along the Th17 pathway [11, 18]. Recent studies have shown that M1 macrophages, in addition to pro-inflammatory properties, exhibit reparative properties: they secrete VEGF, which stimulates angiogenesis and the formation of granulation tissue [21].
Alternative activation of macrophages (M2 phenotype) is observed when they are stimulated by interleukins, glucocorticoids, immune complexes, TLR agonists, etc. They migrate to zones of helminth invasion, accumulate in fibrosis loci, in healing skin wounds and neoplastic formations. M2 macrophages are capable of active proliferation in situ. They exhibit a greater ability for phagocytosis compared to M1 macrophages and express a greater number of associated receptors: CD36 – scavenger receptor of apoptotic cells; CD206 – mannose receptor; CD301 – receptor for galactose and N-acetylglucosamine residues; CD163 is a receptor for the hemoglobin-haptoglobin complex. Macrophages of this type are characterized by a low IL-12/IL-10 ratio[11, 22–25].
Alternatively activated macrophages are divided into subtypes: M2a, M2b and M2c. An example of the M2a phenotype of macrophages are cells that accumulate around helminth and protozoan larvae, the allergens of which induce an immune Th2 response, accompanied by the production of IL-4 and IL-13[23]. They do not secrete significant amounts of proinflammatory cytokines and synthesize a special spectrum of chemokines and membrane receptors [11, 17, 22]. It is believed that they are characterized by the synthesis of IL-10[13], however, in vitro, macrophages do not always produce this cytokine and can exhibit high transcriptional activity of the IL-12 and IL-6 genes[17, 26]. An important characteristic of this population is the synthesis of IL-1 receptor antagonist (IL-1ra), which, by binding to IL-1, blocks its proinflammatory effects[27].
M2a macrophages suppress the inflammatory response by blocking the formation of the M1 population through the cytokines of the Th2 lymphocytes recruited by them, or due to the chemokine CCL17 produced, which, together with IL-10, inhibits the differentiation of macrophages in the M1 direction [11]. M2a phenotype cells are considered typical reparative macrophages. The chemokine CCL2 synthesized by them is a chemoattractant of myofibroblast precursors – fibrocytes[28, 29], they secrete factors that ensure connective tissue remodeling[20, 22, 24, 30–32].
Polarization in the M2b direction is accomplished by stimulation of the Fcg receptor together with TLR agonists and ligands for the IL-1 receptor. Functionally, they are close to M1 macrophages, producing pro-inflammatory mediators and nitrogen monoxide (NO), but at the same time they are characterized by a high level of IL-10 synthesis and reduced IL-12 production [16, 17]. M2b macrophages increase antibody production. The chemokine CCL1 synthesized by them promotes the polarization of lymphocytes in the Th2 direction [11, 13]. M2c macrophages have suppressive properties - they inhibit the activation and proliferation of CD4+ lymphocytes caused by antigenic stimulation and promote the elimination of activated T cells [33]. In vitro, the M2c subtype is obtained by stimulating mononuclear phagocytes with glucocorticoids, IL-10, TGF-β, prostaglandin E2, etc. They do not have bactericidal activity, produce a small amount of cytokines, secrete growth factors and some chemokines [7, 13, 17, 21, 31 ]. M2c macrophages express receptors for phagocytosis and many proinflammatory chemokines, which presumably do not serve to excite the corresponding signals, but are traps for proinflammatory mediators, blocking their functions [11].
The nature of macrophage activation is not strictly determined and stable. The possibility of transformation of the M1 phenotype into M2 has been shown with a change in the spectrum of stimulating cytokines and due to efferocytosis. After engulfing apoptotic cells, macrophages sharply reduce the synthesis and secretion of inflammatory mediators CCL2, CCL3, CXCL1, CXCL 2, TNF-a, MG-CSF, IL-1b, IL-8 and greatly increase the production of TGF-b [20, 25, 27] . The reverse transformation of the M2 phenotype to M1 is expected during the development of obesity.
Many authors question the existence in the body of two clearly distinguishable populations of macrophages M1 and M2. A combination of signs of classical and alternative activation is characteristic of macrophages of human skin wounds. Thus, along with the cytokines TNF-a and IL-12 typical for M1 macrophages, they demonstrate the synthesis of M2 macrophage markers: IL-10, CD206, CD163, CD36 and receptors for IL-4 [34]. A type of macrophages different from M1/M2 with pronounced fibrinolytic activity was found in the liver of mice in a model of reversible fibrosis and in human liver tissue with cirrhosis. They express the genes of arginase 1, mannose receptors and IGF, they secrete MMP-9, MMP-12, exhibit a pronounced ability to proliferation and phagocytosis, but do not synthesize IL-10, IL-1ra, TGF-b[35]. A special population of macrophages is formed in the mouse spleen during infection with mycobacteria. They inhibit the proliferation of T lymphocytes and their secretion of both Th1 and Th2 cytokines, stimulating polarization in Th17. direction. Suppressive macrophages have a unique phenotype - they express genes active in M1 macrophages - IL-12, IL-1b, IL-6, TNF-a, iNOS and at the same time genes CD163, IL-10, mannose receptors and other markers of M2 macrophages [19].
These studies clearly show that the macrophage populations formed in natural conditions differ significantly from the M1 and M2 populations obtained in vitro. Perceiving a variety of activating signals, the macrophage responds “on demand”, secreting mediators adequately to changes in the environment, therefore, in each specific case, its own phenotype is formed, sometimes, perhaps even unique.
Macrophage response to foreign material
Contact of macrophages with foreign material, both in the form of small particles and in the form of extensive surfaces, leads to their activation. One of the serious problems in traumatology and orthopedics associated with a reaction to a foreign body is the development of joint instability after endoprosthetics, which, according to some data, is detected in 25–60% of patients in the first years after the operation and does not tend to decrease[36 ].
The surface of orthopedic prostheses wears out with the formation of particles that infiltrate the soft tissue. The chemical properties of the material determine the possibility of opsonization of particles by blood plasma proteins and the type of surface receptors that initiate phagocytosis. Thus, polyethylene, which activates complement, undergoes opsonization and is “recognized” by the complement receptor CR3, while titanium particles are absorbed by the cell through the opsonin-independent receptor MARCO. Phagocytosis of metal particles, synthetic polymers, ceramics, and hydroxyapatite by macrophages triggers the synthesis of proinflammatory mediators and the osteoclastogenesis inducer RANKL. CCL3 secreted by macrophages causes the migration of osteoclasts, and IL-1b, TNF-a, CCL5 and PGE2 stimulate their differentiation and activation. Osteoclasts resorb bone in the prosthetic area, but new bone formation is suppressed, since the corpuscular material inhibits collagen synthesis, inhibits the proliferation and differentiation of osteoblasts and induces their apoptosis [37, 38]. The inflammatory response caused by wear particles is considered the main cause of osteolysis.
Contact of tissues with material that cannot be phagocytosed initiates a cascade of events known as the foreign body response, or tissue reaction. It consists in the adsorption of plasma proteins, the development of an inflammatory response, initially acute, subsequently chronic, the proliferation of myofibroblasts and fibroblasts and the formation of a fibrous capsule that delimits the foreign body from the surrounding tissues. The main cells of persistent inflammation at the material/tissue interface are macrophages; its severity determines the degree of fibrosis in the contact zone. Interest in the study of tissue reactions is associated primarily with the widespread use of synthetic materials in various fields of medicine [12, 39, 40].
Adsorption of blood plasma proteins is the first stage of interaction of implanted materials with body tissues. The chemical composition, free energy, polarity of surface functional groups, and the degree of surface hydrophilicity determine the amount, composition and conformational changes in the bound proteins, which are the matrix for subsequent cell adhesion, including macrophages. The most significant in this regard are fibrinogen, IgG, complement system proteins, vitronectin, fibronectin and albumin.
A layer of fibrinogen quickly forms on almost all foreign materials. On hydrophobic surfaces, fibrinogen forms a monolayer of tightly bound, partially denatured protein, the epitopes of which are open to interaction with cellular receptors. On hydrophilic materials, fibrinogen is more often deposited in the form of a loose multilayer coating, and the outer layers are weakly or practically not denatured, leaving binding sites inaccessible to cellular receptors of macrophages and platelets[41, 42].
Many synthetic polymers have the ability to absorb components of the complement system and activate it with the formation of the C3-convertase complex. The fragments C3a and C5a generated by it are chemoattractants and activators of phagocytes, iC3b acts as a ligand for the cell adhesion receptor. The activation cascade can be triggered via both classical (mediated by adsorbed JgG molecules) and alternative pathways[43]. The latter is initiated by the binding of the C3 component to surfaces bearing functional groups, for example – OH-, causing its hydrolysis. The alternative pathway can also be switched on after the classical pathway or together with it due to the work of the C3 convertase of the classical pathway, which generates fragments of C3b, the triggering factor of the amplification loop, that are fixed on surfaces. However, sorption and even the beginning hydrolysis of C3 do not always lead to the appearance of an amplification signal. For example, C3 is strongly sorbed by polyvinylpyrrolidone, but its proteolysis on this surface is weakly expressed. Fluorinated surfaces, silicone and polystyrene weakly activate complement. For cellular reactions on foreign surfaces, not only the activation of the complement system is important, but the binding of other proteins mediated by its fragments is important.
The role of albumin lies in its ability to bind proteins of the complement system[12]. It does not promote the adhesion of macrophages and, unlike fibrinogen, does not induce their synthesis of TNF-a[44]. Fibronectin and vitronectin, proteins rich in RGD sequences (amino acid regions ARG-GLY-ASP), are usually found on implanted materials.
With regard to vitronectin, it is unknown whether it is adsorbed directly on the surface of the material or is part of the inactivated membrane attack complement complex fixed on it. Its significance for the development of tissue reaction is that it ensures the strongest and longest-lasting adhesion of macrophages. The interaction of macrophages with the substrate is ensured by cellular receptors for integrin proteins (avβ3, a5β1, CR3), rich in RGD sequences (Table). Blockade of macrophage adhesion by soluble RGD mimetics or removal of the CR3 receptor from their surface reduces the intensity of the tissue reaction, reducing the thickness of the forming fibrous capsule [12, 41, 45].
Attached macrophages fuse to form multinucleated cells (giant foreign body cells - GCTC). Inducers of this process are IFNγ, IL-1, IL-2, IL-3, IL-4, IL-13 and GM-CSF, which stimulate the expression of mannose receptors, which play an important role in cell fusion[12]. GKIT function as macrophages - they have the ability to phagocytose, generate oxygen and nitrogen radicals, synthesize cytokines and growth factors. The nature of the synthetic activity of these cells apparently depends on their “age”: at the early stages of the development of the tissue reaction, IL-1a, TNF-a are expressed, and later there is a switch to anti-inflammatory and profibrogenic mediators - IL-4, IL-10, IL-13, TGF-β[46, 47].
The response of macrophages to foreign materials has been studied under various conditions in vitro and in vivo. In in vitro experiments, the intensity of their adhesion on the surface under study and the formation of HCIT, the number of “turned on” genes, the number of synthesized and secreted enzymes, cytokines and chemokines are taken into account. In monocultures of mononuclear phagocytes adhered to various surfaces, it is not their polarization in the M1 and M2 directions that occurs, but the formation of mixed-type macrophages secreting both pro- and anti-inflammatory mediators with a shift towards the latter during long-term cultivation [12, 48, 49] . The absence of a “gold standard” - a stable control material that has proven itself when implanted into a living organism, with which the tested materials could be compared, as well as the use of non-standardized macrophage cell lines, different methods of their differentiation make it difficult to compare the results of the work of different authors. However, in vitro studies make it possible to judge the cytotoxicity of materials and determine the reaction of macrophages to their chemical modification. Valuable information was obtained by studying the activation of macrophages on the surface of various collagens - native and chemically modified. Native collagens induce in vitro the synthesis of signaling molecules by macrophages, both stimulating the inflammatory response (TNF-a, IL-6, IL-8, IL-1β, IL-12, CCL2) and suppressing it (IL-1ra, IL-10 ), as well as matrix metalloproteases and their inhibitors.[48-50]. The pro-inflammatory properties of such materials depend on the method of decellularization and sterilization of the starting material, which significantly changes its characteristics. Collagen endoprostheses obtained using different technologies from native collagen vary in their ability to induce the expression of proinflammatory cytokines from practically inert to highly active[51]. Injecting collagen with various chemicals changes the nature of the macrophage reaction. Treatment with glutaraldehyde leads to cytotoxicity, manifested in damage to the cytoplasmic membrane, impaired adhesion, and decreased viability of macrophages. At the same time, their production of IL-6 and TNF-a is increased, and the synthesis of IL-1ra is suppressed in comparison with macrophages adhered to native and carbodiimide-stitched collagen. Treatment with carbodiimide provides optimal properties to collagen, which is not cytotoxic, does not cause a significant increase in the secretion of proinflammatory cytokines and metalloproteases, and does not suppress the synthesis of IL-10 and IL-1ra in comparison with native [49].
In order to reduce the tissue reaction, components of the intercellular matrix, native or modified, are introduced into collagen materials. J. Kajahn et al. (2012) created an in vitro imitation of the pro-inflammatory microenvironment of endoprostheses, which promoted the differentiation of monocytes in the M1 direction[50]. Under the same conditions, additional sulfated hyaluronic acid introduced into the collagen substrate reduced the secretion of proinflammatory cytokines by macrophages and increased the production of IL-10. According to the authors, this indicates M2 polarization of macrophages, promoting regeneration and restoration of the functional properties of surrounding tissues. The response of macrophages to slowly degradable and stable materials in vitro is generally uniform and similar to the response to biomaterials, although some specificity of the response is still noticeable. Titanium, polyurethane, polymethyl methacrylate, polytetrafluoroethylene are weak inducers of inflammatory mediators, although titanium promotes higher secretion of TNF-a and IL-10 than polyurethane, and the peculiarity of polypropylene is to stimulate the production of the profibrogenic chemokine CCL18 [52-54]. PEG, proposed as a substrate for cell transfer, causes a sharp but rapidly increasing expression of IL-1β, TNF-a, IL-12, however, its copolymerization with cell adhesion oligopeptide improves the biocompatibility of the material, significantly reducing the expression of pro-inflammatory cytokines[55 ].
The response of macrophages to various materials in vitro does not fully characterize their behavior in the body. In monocultures, there are no factors of interaction with other cell populations and phenotypic polymorphism is not taken into account - in natural conditions, not only monocytic precursors migrate to the implant, but also mature tissue macrophages, the response of which can differ significantly from those recruited from the blood. The study of the secretory activity of macrophages surrounding endoprostheses installed in animal and human tissue is very difficult. The main method for characterizing macrophages based on the M1-M2 paradigm in situ was data from immunocytochemistry of the marker proteins iNOS, CD206, CD163, CD80, CD86. It is postulated that the presence of these markers in macrophages in vivo determines their polarization in the M1 and M2 directions with the synthesis of the corresponding spectra of cyto- and chemokines, but, given the possibility of the existence of mixed type macrophages [34], this characteristic is not entirely correct.
However, in vivo experiments make it possible to trace the fate of the implanted material and the dynamics of the macrophage response over a long period, which is especially important for life-long endoprostheses and devices. The most studied in this aspect are degradable biomaterials based on collagen. The first inflammatory cells to migrate to such materials are PMNs, but this effect is transient and the second wave population is represented by macrophages[56, 57]. Their reaction depends on the physicochemical properties of collagen. The harsher the chemical treatment, the more the collagen differs from the native one, the more “foreign” it becomes for the macrophage and the more pronounced the tissue reaction. Fragments of implants made of slowly degrading stitched collagen installed between the muscle layers of the abdominal wall of a rat promote the formation of GCI and material encapsulation. Migrating macrophages, judging by the expression of the CCR7 and CD206 receptors, can be attributed in some cases to the M1 phenotype, but in many cases it is not possible to determine their belonging to the known phenotypes.
Over time, M2 macrophages appear around the implant, which are located predominantly in the fibrous capsule[58]. Endoprostheses made from non-stitched pig, human and bovine collagen[58] and diisocyanate-stitched sheep collagen[59], which are quickly destroyed in the rat’s body, stimulate the new formation of full-fledged connective and muscle tissue. They do not contribute to the formation of HCIT and are not encapsulated. Some mononuclear phagocytes accumulating at the tissue/material interface do not have M1/M2 phenotype markers, some contain both markers, and some are M2 macrophages. The M1 macrophage subpopulation is absent on such implants[58–60]. Histomorphometric analysis showed a positive correlation between the number of macrophages carrying markers of the M2 phenotype in the early stages of the developing tissue reaction and indicators of successful tissue remodeling in the implantation zone[58].
The tissue reaction to non-degradable materials exists throughout the entire time of their presence in the body[61]. Its intensity is modulated by the physicochemical properties of the materials: in the series polyester, polytetrafluoroethylene, polypropylene - the first polymer causes the most pronounced inflammation and fusion of macrophages, the last - minimal, and the severity of fibrosis for all of these materials positively correlates with the amount of HCIT on the surface of synthetic polymers[40] . Despite the large number of studies that have studied the inflammatory response to various materials, the characteristics of macrophages accumulating on them have not been sufficiently studied. M. T. Wolf et al. (2014) showed that predominantly macrophages with markers of the M1 phenotype (CD86+CD206-) accumulate on the threads and between the nodes of a polypropylene mesh implanted into the abdominal wall of a rat [57].
A gel from the intercellular matrix of connective tissue applied to polypropylene reduces the number of M1 macrophages and GCT and at the same time inhibits the growth of microvessels. This phenomenon is in good agreement with the results of studies demonstrating the expression of M1 angiogenic factors by wound macrophages and the suppression of vasculogenesis during their blockade [21, 52]. Little is known about the synthetic activity of macrophages and the spectrum of their biologically active molecules that provide tissue reactions. In a mouse, macrophages secreting IL-6 and CCL2, IL-13 and TGF-β accumulate at the periphery of the nylon mesh implantation zone, and at the same time, IL-4 is expressed in the cell population, including in the GCIT, adhered to the fibers of the endoprosthesis , IL-10, IL-13 and TGF-β[47]. IL-4 and IL-13 are powerful profibrogenic mediators; they not only polarize macrophages in the M2a direction, promoting the production of growth factors, but also, through the induction of TGF-β expression by fibroblasts, stimulate their collagen synthesis. IL-10 and CCL2 also have a profibrogenic effect, providing chemotaxis of myofibroblast precursors—fibrocytes[29, 30, 62, 63]. It can be assumed that it is macrophages that create an environment conducive to the development of fibrosis around non-degradable materials.
The formation of fibrous tissue can have both negative and positive effects on patient outcomes. In herniological practice, fibrous tissue transformation associated with the implantation of a polypropylene endoprosthesis is one of the main problems (Fig. 2, own data), which, against the background of irrational surgical tactics, in 15–20% of cases leads to the development of recurrent hernias of various localizations [39, 64 ].
In recent years, dental implantation technologies based on the integration of installed structures through the development of connective tissue have been developing especially intensively (Fig. 3, own data). Despite the fact that fibrointegration of implants is recognized by a number of specialists as a valid option, the search for new materials that promote osseointegration processes continues[65].
In this regard, the study of cell populations in the prosthetic area, the development of methods and approaches to blocking an excessive inflammatory reaction leading to fibrosis and stimulating reparative regeneration at the site of implantation of various materials are of significant importance.
Conclusion
Macrophages are a polymorphic population of cells whose phenotype is determined by microenvironmental signals. They play a decisive role in the body’s response to foreign material used for endoprosthetics, catheterization, stenting and other types of treatment. The nature of the reaction and the degree of its severity depend both on the size of the implanted material and on its physicochemical properties and can have both positive and negative implications for the patient’s body. For degradable collagen-based materials, the dependence of the type of macrophage activation and the rate of connective tissue regeneration on the method of processing collagen raw materials has been shown. This opens up great opportunities for specialists developing new methods for tissue decellularization, chemical modification and sterilization of collagen materials in order to obtain implants for regenerative medicine.
Problems associated with the activation of macrophages by non-degradable materials, apparently, should be solved differently. Macrophages phagocytizing wear microparticles on the surface of joint endoprostheses and macrophages migrating to the extensive surfaces of synthetic implants initiate long-term persistent inflammation, osteolysis in the first case and fibrosis in the second. Mitigation of this effect will most likely be achieved by blocking directional migration, adhesion and activation of monocytes/macrophages, which will require deeper knowledge of these processes than we currently have.
Why is an increase in monocytes dangerous?
An increase in monocytes in the body in most cases indicates the presence of pathology. When an infectious disease appears, the body begins a stubborn fight against it by additionally producing white cells to actively fight the disease. It seems that the more monocytes, the better, because this way you can quickly cope with the disease. In fact, there is a downside, since an excessive amount of them can itself provoke inflammation.
Large accumulations of white bodies in the vessels can:
- Disrupt blood flow.
- Strengthen atherosclerosis.
- Reduce blood flow to the heart muscle.
- Cause damage to the vessel wall.